

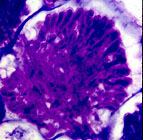
Myrica pensylvanica in sand dunes in New Jersey (top), young bayberry root nodule and vesicle cluster stained with toluidine blue (D. Benson)
Myricaceae Species |
||
| Comptonia peregrina L. (Coult.) Morella californica (Cham. & Schlecht.) Wilbur |
Morella cerifera (L.) Small M. faya (Ait.) Wilbur M. pensylvanica (Mirbel) Kartesz, comb. nov. ined. |
Myrica gale L. M. hartwegii S. Wats. |
(Ecology of the Myricaceae symbioses)
The family Myricaceae has been revised in recent years. In particular, many of the species previously in the genus Myrica have been renamed in the genus Morella. Thus, the family now consists of three genera: Morella, Myrica, and Comptonia with an additional "relictual" genus known as Canacomyrica. The two "myrica" genera are Myrica gale and M. hartwegii, while the majority of former myricas from North and South America, Asia and Africa now come under the genus Morella. Comptonia peregrina L. (Coult.) from North America retains its name.
The family is classified in the Higher Hamamelidae, in the order Fagales (APG, 1998). Molecular phylogenetic studies have not provided strong support for its relationships within the order (Manos and Steele, 1997). Morella is by far the largest genus, having about 50 described species with a wide distribution in North America, Europe, Africa, and Asia. Comptonia peregrina L. is native to North America and Canacomyrica monticola Guillaumin. is endemic to New Caledonia. Nodulation has been observed on all species of Myrica, Morella and Comptonia, but has yet to be documented on Canacomyrica (Navarro et al., 1999). Many Morella spp. have been transported to new sites. For example M. faya has been introduced to Hawaii where it has become an invasive exotic pest (Mabberely, 1988).
To date, there has been no family-wide molecular phylogeny generated for the Myricaceae, yet the fossil history of the family has been well-discussed (see Macdonald, 1989). Briefly, there are two different opinions concerning the first appearance of the Myricaceae in the fossil record. One view holds that the Myricaceae appeared early, during the Santonian period (83–85 mybp), based on Myrica-like pollen. The other view holds that the Myricaceae is instead a much later lineage, originating during the Eocene where there is more fossil evidence, and that the previous fossilized pollen was misidentified (Macdonald, 1989).
The geographic origin of the family is also in dispute. Both a Southeast Asian origin during the early Cretaceous (146 – 65 mybp) and a northern Tethyan origin during the late Cretaceous have been suggested. The enigmatic genus Canacomyrica appears to have many morphological similarities to the fossil, ancestral Juglandaceae and may represent the extant relictual ancestor to the Myricaceae (Macdonald, 1989). If Canacomyrica does indeed represent the basal lineage to the Myricaceae, than it would appear that the ability to nodulate occurred after the family had diverged or that the lineage leading to Canacomyrica lost the ability to nodulate. A phylogeny and detailed biogeographic study is needed to further explore this issue.
For a discussion of the ecological associations between alders and Frankia go to the the Frankia Ecology page. For a discussion of the evolutionary relationships between plant and Frankia go to the Frankia Phylogeny and Plant Phylogeny pages.