
Bayberry root nodule

Myrica pensylvanica in sand dunes in New Jersey
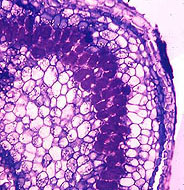
nodule cross-section stained with toluidine blue (D. Benson)
Plant Link - Myricaceae
Members of the Myricaceae, Morella, Comptonia and Myrica sp., have been considered to be promiscuous hosts based on the results of greenhouse cross-inoculation studies and on ecological evidence (Baker, 1987; Clawson et al., 1998; Torrey, 1990). These plants are nodulated by frankiae from phylogenetic Clusters 1 and 3. Cluster 2 strains have not been detected in the family.
The natural diversity of Frankia strains in the nodules of Myrica (Morella) spp. native to the Northeast North America has been examined using the variability between 16S rDNA sequences or PCR-RFLP of 16S rDNA as measures of diversity (Clawson and Benson, 1999; Huguet et al., 2001). In one study, root nodules were collected from 30 sites with Comptonia peregrina, 29 with Myrica pensylvanica and 37 with M. gale. Thirty-seven unique sequences were found in 97 nodules analyzed. Only two were present in all three plant species and two more were found in both C. peregrina and M. pensylvanica.
Interestingly, the richness and evenness components of diversity differed markedly between plant species. Nine different Cluster 1 sequences were obtained from 37 M. gale nodules with three accounting for 81% of the total. C. peregrina nodules had 15 Cluster 1 sequences in 30 nodules with four accounting for 60% of the total. Bayberry (Morella pensylvanica) nodules yielded the highest diversity with 20 Frankia strain sequences in 29 nodules. Of the twenty sequences, thirteen from Cluster 1 were found in twenty nodules, six Cluster 3 sequences were found in eight nodules, and one nodule yielded a sequence like that of Nod-/Fix- actinomycetes isolated from a variety of actinorhizal plants (Clawson, et al., 1998). Identical sequences were commonly found in plants growing at widely dispersed sites indicating that some Frankia strains are cosmopolitan.
The case of M. gale is especially interesting. It had been considered to be a “promiscuous host” since it nodulated with most Frankia isolates in greenhouse trials (Torrey, 1990). It is less promiscuous in nature. This observation has been further confirmed in a separate study using PCR-RFLP of 16S rDNA PCR amplified from root nodules collected from M. gale nodules in Canada (Huguet, et al., 2001). Low diversity in M. gale nodules may be attributed to its preference for growing in water-saturated soils near lakes, swamps or bogs. Such locations are typically acidic and low in oxygen; such conditions may limit the selection of Frankia to those strains capable of tolerating them.
In older studies, M. gale exhibited distinctive patterns of distribution of sp(+) and sp(-) (indicating the presence or absence of Frankia sporangia in nodules) root nodules with sp(+) strains more common in nodules collected at southern interior and coastal regions and the sp(-) strains more common in northern and western Maine (Schwintzer, 1990). The presence of sp(+) nodules positively correlated with the average number of frost-free days per year, and with the % organic matter in the soil, and negatively correlated with pH, with more sp(+) nodules found in more acidic soils. Although the sporulation phenotype has low resolution, its significance is enhanced by the finding that the diversity of frankiae in these nodules is low. A similar sorting by environment has been observed in sp(+) and sp(-) nodules growing on alders (see above). Thus, local edaphic factors are clearly important in determining which Frankia strains get into nodules and most likely how strains distribute among environments.
Since Myrica spp. and C. peregrina are nodulated by "alder strains" from Cluster 1 and "elaeagnus strains" from Cluster 3, the environmental distribution of strains that nodulate myricas is very wide.