
Casuarina cunninghamiana in the greenhouse (D. Benson).

C. cunninghamiana crown nodule (D. Benson)
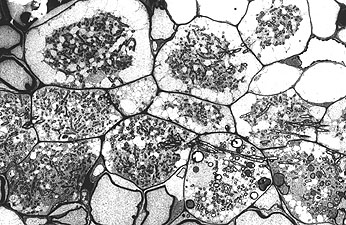
TEM of infected cells (R. H. Berg)
Plant link - Casuarinaceae
Although the Casuarinaceae is sister to the Betulaceae and Myricaceae among the higher hamamelids, and all three families are infected by Cluster 1 frankiae, casuarina strains are not ubiquitous in soils (Diem and Dommergues, 1990). For example, in Jamaica, Myrica strains were abundant in all soils tested but no casuarina strains were detected using C. cunninghamiana as the test plant outside of casuarina stands (Zimpfer et al., 1997). C. cunninghamiana is however nodulated where it has been established in Jamaica suggesting that a strain had been introduced along with the plant but has not spread. In soil collected along transects leading away from C. cunninghamiana trees, casuarina frankiae were undetectable at about 20 m away from the trees. On the other hand Myrica frankiae were found at all distances despite the absence of a host (Zimpfer, et al., 1999). This observation may account for the fact that Casuarina plants must be inoculated in most areas of the world (Diem and Dommergues, 1990; Simonet, et al., 1999).
It has become apparent that a narrow group of closely related strains is responsible for nodulating Casuarina and Allocasuarina spp. outside their normal geographic range (Fernandez, et al., 1989; Honerlage et al., 1994; Maggia et al., 1992; Nazaret et al., 1991; Nazaret et al., 1989; Rouvier et al., 1990; Rouvier, et al., 1996; Simonet, et al., 1999). In a study that looked at casuarina frankiae in nodules collected from around the world, all but one nodule from five Casuarina and Allocasuarina species yielded the same PCR-RFLP group that dominates in culture collections. This group was delineated using DNA-DNA reassociation (Fernandez, et al., 1989), and by PCR-RFLP of the intergenic regions between the rrs and rrl genes and between the nifH and nifD genes (Honerlage, et al., 1994; Maggia, et al., 1992; Rouvier et al., 1992; Simonet, et al., 1999). Several members of this group have had their genomes sequenced (Belaid et al., 2020).
Casuarina frankiae diversity is greater within the native range of the plants. For example, 22 nodules collected from two Casuarina spp. and two Allocasuarina spp. in Australia yielded a total of five rrn and nif PCR-RFLP groups (Rouvier, et al., 1996), although the dominant strains were still in the globally dispersed PCR-RFLP group. Despite the relatively small number of nodules used in the study a degree of plant species-Frankia strain specificity was noted. In another study, seven groups were identified among 110 nodules sampled from five Casuarinaceae species in Australia (Simonet, et al., 1999). Each IGS group seemed limited to one or two of the plant species. This specificity could be due to the host plant selecting specific strains in the rhizosphere from among a population of strains, or more likely to environmental selection limiting the types of frankiae that are available in a particular soil.
In contrast to its nearest relatives in the Casuarinaceae, Gymnostoma spp. are nodulated by members of Cluster 3 Frankia strains rather than by strains from Cluster 1. The reason for this symbiont shift may be related to an early allopatric distribution of the plants and presumably symbionts (Gymnostoma in the north and east of Gondwana islands; Casuarina/Allocasuarina in the drier Australia) (Simonet, et al., 1999). Consistent with this hypothesis is the observation that the only Casuarina species present in New Caledonia, C. collina, commonly contains both Cluster 1 casuarina strains and Cluster 3 strains; the latter are similar or identical to those nodulating Gymnostoma, also found on the island (Gauthier et al., 1999).
A study of Frankia diversity in Gymnostoma nodules collected from New Caledonia reported 17 different RFLP patterns from 358 nodules collected from eight Gymnostoma species (Navarro, et al., 1999). Species specificity was not noted but a correlation was found between soil type, host species and RFLP pattern. A dominant pattern accounted for 56% of the total but was absent from 45 nodules obtained from plants growing at acidic sites and was thus deemed specific for ultramafic soils. Another pattern belonged to a more cosmopolitan strain group found in all species on all soils and was represented in 14.5% of the total. Strains that nodulate Gymnostoma are wide-spread in New Caledonia with populations being amplified in soils beneath Gymnostoma (Gauthier et al., 2000), and in the rhizosphere of a member of a non-nodulating member of the Rhamnaceae (Gauthier, et al., 2000).
The conclusion from these studies and others (Jaffre et al., 2001) is that different populations of Cluster 3 frankiae colonize different Gymnostoma spp. more as a function of soil type rather than species specificity. This work, together with M. gale work (Clawson and Benson, 1999a; Huguet et al, 2001) and Alnus incana work (Clawson and Benson 1999b), provide the strongest examples of strain sorting by environment.