
Ceanothus velutinus (light green) regrowing after a fire in the Sierra Nevada mountains.

Colletia hystrix (top right) regrowing after fire in the Andes mountains, Chile (D. Benson)
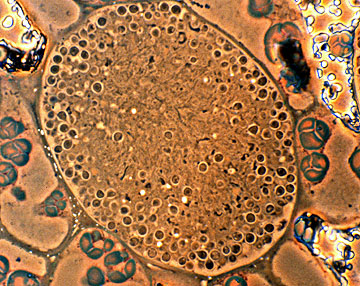
Vesicle cluster in root nodules of Ceanothus americanus (R. H. Berg)
Plant link - (The Rhamnaceae)
The Rhamnaceae frankiae provide something of a conundrum. Only six out of 50 plant genera have members that are nodulated. Five of the genera are within the tribe Colletieae and are located in the southern hemisphere; they are nodulated by Cluster 3 ("elaeagnus") Frankia. Several Frankia strains have been isolated from the root nodules of the South American Colletieae (Carú, 1993; Schwencke and Caru, 2001), but the ecological diversity patterns are not yet known.
One genus, Ceanothus, is limited to the northern hemisphere, from the western North America from Canada to Mexico; it is nodulated by members of the enigmatic Cluster 2 Frankia that are similar or identical to those that nodulate Datisca, Coriaria and the actinorhizal Rosaceae (Benson and Clawson, 2000; Vanden Heuvel et al., 2004).
A number of studies have addressed the ecology of Ceanothus frankiae. One study on Ceanothus americanus found relatively high diversity of Frankia strains as assessed with RFLP of total DNAs probed with nifDH genes or with random probes (Baker and Mullin, 1994). Another used repetitive extragenic palindromic PCR (Rep-PCR) and found 11 patterns in 54 Ceanothus spp. nodules from seven sites in southern California (Murry, et al., 1997). 16S rRNA gene sequencing from the latter study indicated habitation by Cluster 3 Frankia strains, that is, elaeagnus strains. In addition, some isolates from Ceanothus nodules infect Elaeagnus spp. but cannot reinfect Ceanothus plants (Lechevalier and Ruan, 1984; Ramirez-Saad et al., 1998; Torrey, 1990). More recent studies that have removed nodule periderms before preparing template DNA for PCR amplification have found only Cluster 2 strains in Ceanothus nodules (Clawson et al., 1996; Vanden Heuvel et al., 2004).
A relationship between strains and the soil conditions from where nodules were harvested has been observed using PCR-RFLP analysis of the ribosomal rrs-rrl region (Ritchie and Myrold, 1999). Of four RFLP groups identified, one occured in mountainous areas and two others were limited to the Willamette Valley. The fourth group was limited to C. americanus collected from Tennessee. Overall, the diversity of strains reported was less than that reported using other methods. In a similar manner, sampling of nodules from co-populations of different Ceanothus species indicated that Frankia strain PCR-RFLP patterns were more likely to be related to the environment from which the nodules came than to the plant species infected (Jeong, 2001; Jeong and Myrold, 1999).
Ceanothus frankiae have been enumerated by trapping experiments from soils. Populations are amplified beneath Ceanothus stands although sites without host plants retained a small population (Jeong, 2001; Wollum II, et al., 1968). Even in soil beneath host plants, the nodulation capacity is low and estimated at 3.6 to 5.2 NUs x g-1 soil (Jeong, 2001). Such a low apparent population density may either be typical of Ceanothus strains or may reflect the difficulty nodulating Ceanothus plants in the greenhouse (Rojas et al., 2001).
Ribosomal RNA gene sequences amplified from Ceanothus nodules are generally very similar (99-100%) to each other and to those found in the Rosaceae, Datiscaceae and Coriariaceae. This low diversity suggests that some Cluster 2 strains are globally dominant (Benson, et al., 1996; Ramirez-Saad, et al., 1998; Ritchie and Myrold, 1999). Plants from these families are all found in western North America although Coriaria and Datisca have disjunct populations in several parts of the world (Benson and Clawson, 2000). It is possible that Ceanothus became geographically isolated from other Rhamnaceae and subsequently specialized in the Cluster 2 Frankia strains that may have been more adapted to the environment or simply more numerous because of their proximity to other actinorhizal plants.